
[HOME PAGE] [CURRICULUM VITAE] [DRAFTS AND SKETCHES]
23/01/2003 - http://forums.about.com/ab-christianity/messages?msg=9881.141
Darwinia passed on to me some of the specious arguments people are still spouting in an attempt to refute the theory of evolution. It took me about half an hour to dig out the attached illustrations plus another hour or two mucking about with the scanner to get good images that don't take up too much space - and a bit more time to put the words around them -- and then a day to work out how to get the graphics into the forum.
If I had the time and inclination, I can put up similar cases against any of the other arguments from design creationists have tried to support. Genetics, population biology and natural selection provide an entirely non-mystical explanation for the diversity of life as it exists today. Those who argue the explanations are inadequate simply lack the knowledge of what exists in the world today. Anyway, following is the essay Darwinia referred to:
As someone who has taught both vertebrate and invertebrate biology at the university level in the late 60's and 70's from evolutionary points of view, I become very frustrated with ignorant people who argue that processes of random variation guided by natural selection could not possibly lead to the natural evolution of organs as complex as the vertebrate eye. They argue that there is no possibility of intermediate structures that would in themselves be adaptive to the organisms possessing them, therefore such marvellous structures must have been "engineered" by an intelligent designer.
People who make such unsupportable statements are simply ignorant of the diversity of living organisms. The reality of living things refutes their arguments. Taking the vertebrate (i.e., human) eye as a commonly cited example, the fact is that one can find almost every possible step in the evolution of this complex organ in living organisms today. "Intermediates" range from single celled flagellated protozoa able to orient to light, to highly complex organisms such as insects, squid and people.
Following are some illustrations of functionally successful structural intermediates along the line of development of a "camera" type of eye collected from some of the reference materials and textbooks I used as texts or background for my lectures and lab exercises. Comparable intermediate stages may be found in the evolution of the compound eye typical of insects and crustacea (spiders do a bit of both - they generally have 8 eyes ranging from camera type to photosensors not unlike single elements comprising the compound eye). And as I said, the same applies for virtually any other organ (including the human brain) creationists throw up as an example of something that couldn't have evolved through natural (as opposed to mystical) processes.
As a caveat, it should be recognised that the examples given below do NOT represent a genetic or historical evolutionary sequence from primitive to advanced, as each species in the structural series is living today and has visual capabilities apparently sufficient to their needs (i.e., there is a considerable developmental cost and no advantage to having an image forming eye if the nervous system doesn't have the capability to process the image and make decisions relating to it). However, it is easy to consider that changed circumstances would lead to selective pressures to "advance" or "regress" from any point along the scale of examples given.
Ignorance of the diversity of life is understandable. Most structural evolution has taken place in marine habitats in small to microscopic organisms (and especially their larval stages) that would be unfamiliar even to biologists studying life on the land or fossils. I grew up on a boat, and spent a significant part of my childhood immersed in Catalina Island tide pools. I have also had the good fortune to teach in areas where it was possible to sample the diversity of marine organisms and maintain living examples in marine aquaria where I could mount stereo microscopes to look into the side of the aquarium and see with my own senses the world from the point of view of the microscopic and near microscopic marine organisms.
All that is required to see many examples of most of the marine phyla in vivo is a tow of a plankton net and to collect some seaweed, a few mossy rocks and some bottom sand and mud into the aquarium and begin watching the animal life that comes with it. Even tinier organisms may be seen and studied in life with a long-focus compound microscope. If phase contrast optics are available, this is even better.
My series of examples begins with single celled flagellated protozoa such as Euglena that respond to light to regulate their rate of photosynthesis. This is achieved through a stigma (or "eye spot") that shades the base of a flagellum and thus controls the type of motion it exhibits. I have spent many hours watching such single celled organisms under the microscope and have performed my own simple experiments to demonstrate their photosensitivity and their ability to orient to light. Even single cells with no flagella such amebas can be shown to be light sensitive. Placed in a small petri dish or in the well of a microscope slide they will move as fast as they can from an area exposed to direct light to one in shade (a common lab experiment).
The above illustrations are from Fig. 34 page 104 from Kudo, R.R. 1966. Protozoology, 5th Ed. Charles C. Thomas, Springfield, Ill.
a. is Trachelomonas hispida x 530. b and c are living and stained reproductive cells of Pleodorina illinoisensis x 1000 (Pleodorina is a colonial flagellate that is simpler than the more familiar Volvox - its colonies are flat rather than spherical like Volvox). "Chromatophores" are chloroplasts.
Flatworms also orient to the intensity and direction of light (i.e., they will seek out darker areas as can be demonstrated with living organisms in the lab or nature). The cup eyes give more information about the specific direction the light comes from, and they are also able to move a lot faster than the single celled amebas.
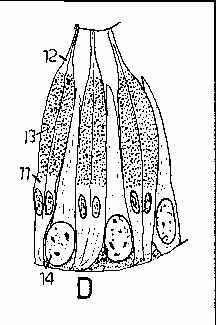
The next three illustrations are flatworm eyespots from Fig 10, page 38 of Hyman, L.H. 1951. The Invertebrates, vol 2 - Platyhelminthes and Rhynchocoela The acoelomate Bilateria. McGraw-Hill Book Company, New York.

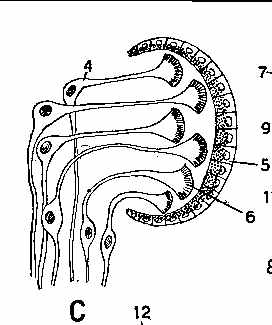

In multicellular organisms, the simplest photosensory organ (B, above) simply consists of
photosensitive cells located in the epidermis that are hooked into some kind of nerve net able to
control the speed and direction of orientation. It doesn't need a "brain". Many lab robots
have been built using simple neural networks that will show similar behaviors to flatworms. In a
slightly more complex structure organising the photoreceptor cells in a partially shaded cup gives
more information regarding the direction the light is coming from. C, above, shows the layout of a
planarian eyecup, where the retinal cells point away from the light (as is the case in humans and
other vertebrates). Planaria is (or at least was when I was teaching) a common experimental organism
for use in freshman biology labs. These have these kinds of eye cups. D, above, shows a more
complex eye from a marine flatworm containing a lens, where the retinal cells point towards the
light source (as is the case in squid and octopus eyes). In the diagrams 5 points to pigment cells,
4 and 6 to photosensitive rod cells, 12 is a transparent "cornea", 13 is a lens and 14 is
a vitreous body.
Larger and more complex cups can begin to function as pinhole cameras, actually able to form fuzzy images of potentially threatening shadows. The next 2 illustrations are from Fig 121 on page 270 of Hyman, L.H. 1967. The Invertebrates, Volume VI, Mollusca I - Aplacophora, Polyplacophora, Monoplacophora, Gastropoda - The coelomate Bilateria.
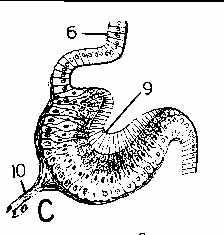
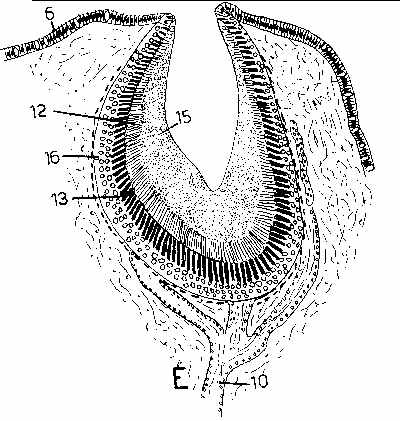
C, above, is an eyespot of a limpet. D, above, is a magnified view of the retina of C. E, above, is from another kind of limpet. In the figures, 6 is the epidermis, 9 is the optic cup, 10 is the optic nerve connecting to the phtotosensitive retinal cells. 11 are pigmented cells 12 are the clear distal ends of pigmented cells. 13 is the pigmented zone. 14 are retinal cells (between the pigment cells in D). 15 is gelatinous material, and 15 is a layer of ganglion cells interposed between the retinal cells and the retinal cells. Note: surrounding the photosensitive cells with pigment ensures that they will respond to light only along their axes - not their sides, so directional information is provided because only those cells fire nerve impulses that are pointed more or less directly towards the light source.
The photosensitive cups can be filled (and the photosensors protected from damage) by transparent secretions and covered over with a transparent epidermis. (Secretion is a normal process for many epidermal cells anyway.) Because the secretions and epidermis itself have a different refractive index than the surrounding water, if they are curved they can work as lenses to bring light to a sharper focus. More exact curves enable sharper focus. The next two pictures are from Fig 94 on page 471 of Hyman (loc cit.) and show eyes of two gastropods where the eyes are at or near the end of tentacles ("intermediate_lens" is from Aplesia, a large sea slug). Note: if you look closely at a garden snail, the black spots at the end of the two longest tentacles are eyes.
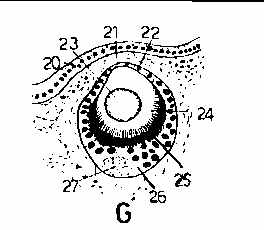
For the callouts in these figures, 20 are epidermal cells, 21 is a blood sinus, 22 is the "cornea", 23 is the lens, 24 is the retinal rod cell border, 25 is a pigment layer, 26 is the retina proper, 27 is the optic nerve, and 28 general connective tissue.
The next to last example is an eye of the pecten, a bivalve mollusc that swims by clapping its shells together. In life, there are a number of eyes in the edge of the mantle that are easily seen as bright blue spots around the edge of the shell. When skin diving I have chased and caught them, and watched them swim in aquaria. The picture is from Figure 8.10 on page 230 of Sherman, I.W. & Sherman V.G. 1970. The Invertebrates: Function and Form: A Laboratory Guide. The Macmillan Company, New York.
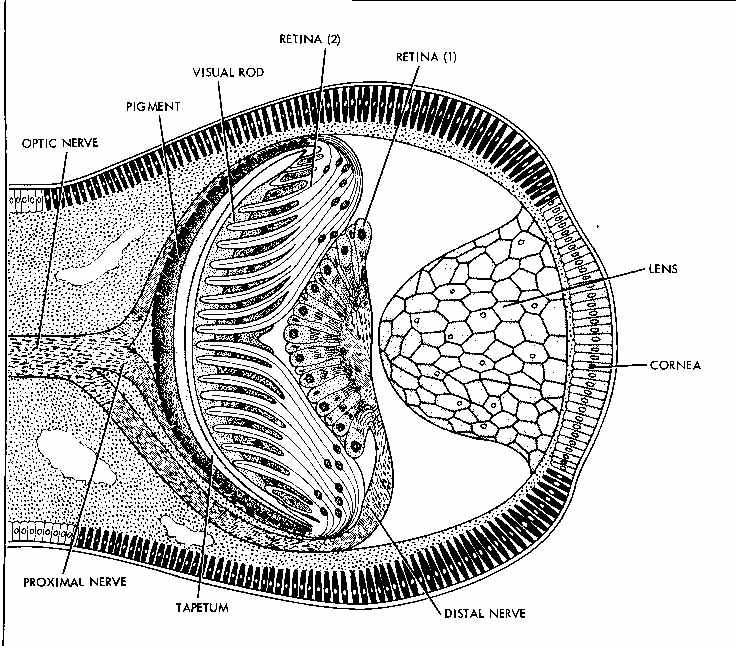
Again, there are easy intermediates to evolve the fully developed camera eye found in the cephalopods, fish and humans. The last picture is from the same figure cited above, showing the eye of a squid, which is at least as sophisticated as the vertebrate eye:

None of the above requires any "intelligent designer" to achieve. Eyes are simply the adaptive responses shaped by natural selection over hundreds of thousands to hundreds of millions of years working on species to meet changing environmental circumstances where single individuals have life spans of weeks to a year, and where single individuals may produce hundreds to millions of larvae with the expectation that on average on of those hundreds to millions will survive to replace the parent.
My bible is the diversity of life as it exists in reality. The story it tells does not require some fantasy creator to make it happen. All that is required to make it happen is a flux of energy and the laws of physics, chemistry and non-equilibrium thermodynamics. This does not answer the question as to how those laws came to exist in the first place, but at least life as it exists on Earth doesn't require or need any mythological father, son and holy ghost to make it all happen.
In all of the diversity of life I have studied and what I know about complex morphologies (i.e., body plans, wings, ears, eyes, etc.) that the creationists and un "intelligent designers" throw up as supposed refutations of evolution and natural selection I have found nothing that cannot reasonably be accounted for by the entirely un-mystical actions of natural selection working on pre-existing materials and processes.